ABSTRACT
Many previous societies
have killed themselves off and, in the process, devastated their
environments. Perhaps the most
famous of these is that of “Easter Island”. This suggests a grand challenge for the
agent community: that of discovering what kinds of rationality and/or
coordination mechanisms would allow humans and the greatest possible variety of
other species to coexist. In particular, solving this challenge consists of
designing and releasing a society of plausible agents into a simulated ecology
and assessing: (a) whether the agents survive and (b) if they do survive, what
impact they have upon the diversity of other species in the simulation. The simulated ecology needs to implement
a suitably dynamic, complex and reactive environment for the test to be
meaningful. Agents, as any other entity have to eat other entities to survive,
but if they destroy the species they depend upon they are likely to die off
themselves. Up to now there has
been a lack of simulations that combine a complex model of the ecology with a
multi-agent model of society. A suitable dynamic ecological model and simple
tests with agents are described to illustrate this challenge.
Categories
and Subject Descriptors
G.3 [Simulation
and Modeling]. J.4 [Social and
Behavioral Sciences]: – sociology. K.4 [Computers and Society]. I.2.11 [Distributed Artificial Intelligence] – multiagent systems. J.3 [Life and Medical Sciences] –
biology and genetics.
General
Terms
Design, Experimentation, Human Factors.
Keywords
Agent-Based Modelling, Ecology, Social Intelligence, Ecological
Intelligence, Resilience, Grand Challenge.
1.
THE
DANGERS OF ECOLOGICAL DAMAGE AND SOCIETAL COLLAPSE
The evidence is overwhelming that, many times,
humans have destroyed the ecologies they inhabited to their own and other
species detriment – sometimes causing whole settlements or civilizations
to disappear. Examples include: the
inhabitants of Easter Island who built its famous stone statues in a race for
status and killed all trees on the island in the process [12], or the Mayan civilization where a combination
of increasing climatic aridity, demands of agriculture and societal conflicts
lead to an abandonment of their impressive step temples in the jungle [14].
However you look at it, humans have a profound effect upon the
ecosystems they come into contact with, even to the extent that (as some have
argued) we are in the middle of the sixth great extinction event – the Holocene [8].
However, how
humans will effect a particular ecosystem is not always clear – sometimes
it seems that a balance between humans and the rest of the ecosystem is
established, but at other times, the arrival of humans can only be described as
catastrophic [7]. The “Social Intelligence Hypothesis” [11] suggests that the main adaptive advantage that
our brains give us is our ability to socially organize. From this view our
brains provide us with social intelligence first (for example abilities to:
recognize other individuals, to develop a personal identity relative to a
group, to be able to communicate, to be sensitive to status, to imitate, to train
our offspring for a long time, and to adsorb a whole culture when experienced
over a long time); any “general” intelligence we have as individuals is a
by-product of these social abilities.
Due to these social
abilities, groups of humans can inhabit a variety of ecological niches. They do
this by adapting to a niche in terms of developing a body of knowledge,
including words, ideas, techniques, social norms, systems of value and ways of organizing,
that enables the group to survive there [13]. Once established, this body of knowledge can
be passed down to new members of the group so that the group can retain its ability
to survive in that niche over time. Broadly, this set of knowledge can be
associated with the culture of the group. Thus, the abilities of groups of
humans can change far more rapidly than that of most animals that have to rely
on genetic evolution. Humans are thus at a distinct advantage in terms of any
adaptive “arms race” with other species. Their social intelligence has equipped
them to survive in a hostile and unpredictable world, ensuring their own
immediate survival as their priority (as with other species).
However, they do not
necessarily plan for the long-term and can cause such a degree of environmental
damage to the niches they inhabit that they endanger the survival of their own
group [7]. In this way, the arrival of humans within a
system of ecosystems can have a profound impact – not merely changing the
extent of extinction but also the whole way that the dynamics of that ecosystem
works. The abilities of humans are over-tuned towards immediate survival, with
the contrary result that, in the longer-term, they grab resources to themselves
in a way that can jeopardize their own group survival.
Now that humans, using their technology, can
inhabit almost any ecological niche on earth, any ecological disaster that we
cause might well not be limited to a particular niche but may affect us globally.
This challenge is thus to use agent-based simulation techniques to help address
this problem, and thus contributing to the survival of our and other species on
this planet. This challenge can be seen as an amplification of that implicitly
posed in [2] or else a contribution to the wider challenge
posed in [7].
2. THE PARTICULAR CHALLENGE FOR THE MULTI-AGENT COMMUNITY
There have been many
agent-based simulations addressing the interaction of man with the environment,
going back (at least) to 1994 [1] (see [3] for a review). Individual-based ecological
models go back even further (see [9] for a review). However to fully address this challenge
we need to have a multi-agent model concerning human decision making and social
interaction combined with an
individual-based model of an ecology that more fully reflects the dynamism and
complexity of real ecologies. Up to
now, models of humans interacting with their environment have had either a
relatively simple model of human interaction or a simple model of the ecosystem
they are embedded in (such as a systems dynamic model). As it said in [6] in 2012:
“…The more serious
shortcomings of existing modelling techniques, however, are of a structural
nature: the failure to adequately capture nonlinear feedbacks within resource
and environmental systems and between human societies and these systems.” (p.
523)
In other words, to
fully address this challenge we need to start to understand how the complexity
of human cognition, the complexity of human society and the complexity of dynamic
ecologies might interact. Otherwise, we might miss some of the complications
that might affect our and other species survival.
To ensure
that the environment in which the agents representing humans and their society
is sufficiently challenging, we require a model of this environment that
satisfies the following criteria.
a) The environment needs to include space, so that
there can be a differentiation in terms of niches and allow for some spatial migration
between parts of the landscape
b) The environment needs to include niches with
different characteristics, for example deserts (which can not sustain life) and
natural barriers (which impede migration)
c) Complex food webs of species need to be able to
develop within each niche either extracting resources from the environment or
other individuals (predation)
d) New species need to be able to evolve in
response to the pressure of the environment, other species and humans
e) Agents representing humans need to be embedded
within these niches, needing to use/eat other species to enable their own
survival
Once such a
test bed is established, the challenge would be implemented in several phases:
1. Bed in
the ecology. Run the ecological model for a while to
allow a rich and dynamic ecology to evolve.
2. ‘Freeze’
the ecology and save this state as the starting point for different evaluative
runs.
3. Inject
the human agents. Then place a
small society of agents with given cognitive and social abilities into the
ecological simulation.
4. Assess
the result. After a suitable
period of simulation time assess the outcome.
The
assessment of the final state of the simulation could be done in a variety of
ways, including:
· Measuring the diversity of the ecology, for
example the average genetic difference between individuals, as in [5] (excluding humans).
· The species-number distribution – how
many species are there with a population of at least 2n, where n varies (the “Species Abundance
Distribution” of [10]).
· The number of trophic layers that have survived
for a period of time since the injection of human agents, shown by the
distribution of trophic layers.
· The health of the society of agents, in terms
of the number of surviving humans and its variability over time.
Measures such as these
can be brought together to assess the sustainability/health of the
socio-ecological system as a whole.
Thus this
challenge can be encapsulated as follows:
To design plausible cognitive and social
abilities that, when implemented in agents and assessed in the above way,
reliably result in a sustainable and healthy socio-ecological system.
3. TOWARDS A CHALLENGE TEST-BED
To illustrate this
challenge I describe a simulation test-bed that meets the stated criteria, has
been tested with simple agents representing humans, and assessed in some of the
above ways.
3.1 The Basic Set-up
In this, entities,
plants, herbivores and predators, are represented as individual objects. They
inhabit one of a number of patches arranged in a 2D grid that makes up the
world. Each patch is well mixed so that interactions within that patch are
random, but there is a probability that each individual can migrate to one of
the four neighbouring patches each tick. Each patch and individual has a binary
bit-string that represents its characteristics. There is a basic energy
economy; so that energy is ‘rained’ down into the world (each tick), divided
equally between patches, and which ultimately drives the whole ecology. These
bit-strings and a fixed random interaction matrix, described below, determine
whether an individual can extract energy from a patch or predate upon another.
The bit-string of any individual is passed to any progeny but there is a
probability that one of the significant bits of their characteristic is flipped
at birth.
3.2 Species Abilities and the Energy
Extraction/Predation they Allow
Key to this understanding
this simulation is how it is determined whether individuals can extract energy
from a patch or predate upon another. This method is adapted from that in [4]. A random interaction matrix with the
dimensions of the length of individuals’ bit-strings is generated at the start
of a simulation. It is filled with normally distributed random floating-point
numbers (mean 0, SD 1/3). This interaction matrix determines which entity can
eat another entity as follows: (1) the non-zero bits of the predator select the
columns of the matrix, the non-zero bits of the potential prey select the rows;
(2) the intersection of the selected rows and columns determine a set of
numbers, (3) these are summed; (4) if the sum is greater than zero the predator
can eat the prey, in which case the prey dies and the predator gains a
percentage of its energy value (the rest is lost). This calculation is
illustrated in Fig. 1.
Essentially the same
process is used to determine which entities can extract energy directly from
the environment, except that the part of the prey is taken by the patch with
its bit string (padded with zeros to reach the appropriate length). In this
case only those with scores greater than zero get any of the patch’s energy.
The patch’s energy is divided between all qualifying individuals in proportion
to their score against the patch. This scheme has the consequence that no individuals
can extract energy from a patch with a bit-string of all zeros (a ‘dessert’).
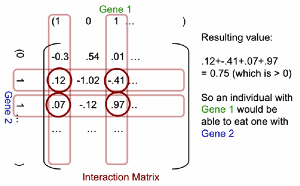
Fig. 1.
The use of the interaction matrix to determine predation as well as
energy extraction from a patch to give its relative fitness
This interaction
scheme allows complex food webs to be evolved, for example via a genetic
“arms-race” between predator species and prey species, since it allows for
adaption with respect to another specific species. It also allows for
competitive adaption to particular kinds of patches. In other words fitness is not an
absolute number but relative to the environment and the other existing species,
if it extracts energy from this, or another species. [4] showed that this kind of scheme can be used to
evolve complex ecologies with plausible characteristics including food webs
with similar network characteristics to those of observed food-webs (however
this was for a single patch).
3.3 Simulation Execution
At the start of the
simulation, the random interaction matrix is generated. Each patch is allocated
a random bit-string with the given number of bits, padded out with zeros to
make it the same length as individuals’ bit-strings. The “environmental
complexity” is the number of significant characteristics that patches can have
– the number of bits in their bit-string. Bit strings of length 2 allow
for 4 types of patch, of length 3 8 types etc.
The
simulation starts with no individuals. Each tick:
· Energy
Distribution. A fixed amount
of energy is added to the model, equally divided between all the patches.
· Death. A life tax is subtracted from all
individuals, if their total energy is less than zero it is removed.
· Initial
seeding. (In the initial phase),
until a viable population is established, a single random individual is
introduced with a given probability each tick.
· Energy
extraction from patch. The
energy stored in a patch is divided among the individuals on that patch that
have a positive score when its bit-string is evaluated against the patch’s
bit-string (in the above manner) in proportion to its relative fitness, at the
simulation’s efficiency rate.
· Predation. In a random order, each individual is
randomly paired with a given number of others on the patch. If it has a
positive dominance score against the other, the other is removed and the
individual gains a fixed proportion of its energy, given by the “efficiency”
parameter.
· Maximum
Store. Individuals can only
retain so much energy, so any above the maximum level set is discarded.
· Birth. If an entity has a level of energy > the “reproduce-level”,
it gives birth to a new entity with the same bit-string as itself, with a
probability of mutation. The new entity has an energy of 1, subtracted from the
energy of the parent.
· Migration. With a probability determined by the
“migration” parameter, the individual is moved to one of the neighbouring 4
patches.
3.4 The ‘Human’ Agents
Broadly speaking, the
agents representing humans should be processed in manner similar to any other
individual with only a few differences.
The most important difference is in the acquisition and passing on of
techniques among their own group.
Thus their “bit string” that determines their ability to predate upon
(or resist being predated upon) is not determined genetically but can be
learned socially by imitation from parents and/or peers. Whether an agent
predates upon another individual and when it moves to a neighbouring patch
could be part of what is determined by the agent’s decision processes. One might well set the required minimum
energy that humans need to give birth as much higher than for other individuals
and allow them to store more energy. They might have a complex social structure
with food passing between themselves according to its rules (e.g. an internal
economy). They may have a tribal
structure that allows each individual to recognise others from their own tribe
and those who are outsiders, which may affect their behaviour. Many other extensions are possible to
reflect other human attributes, e.g. warfare between groups, deliberate
planting of crops or hoarding.
3.5 Some Illustrative Results
Figure 1 shows the a
graph of the number of species in a typical initial stage of the model, showing
the development of plants, then herbivores and finally predators, providing a
suitably complex and dynamic environment, with a range of trophic levels, ready
for the injection of agents representing humans.
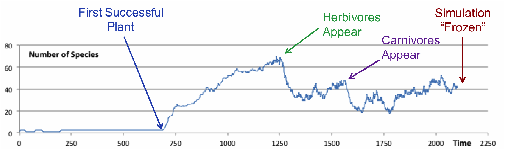
Figure 2. A typical run of the model during the “Bedding In” phase, making the
state of the simulation suitable for the injection of agents representing
humans
To give a simple
flavour of some of the possible results, some very simple agents were injected
at the point indicated in Fig. 2.
Then the simulation was run for a further 1000 simulation ticks with
different migration rates (the probability any entity or agent would move to a
nearby patch in the 2D grid). 25
otherwise independent runs were performed both with and without ‘human’ agents
added, and the final mean ecological diversity measured.
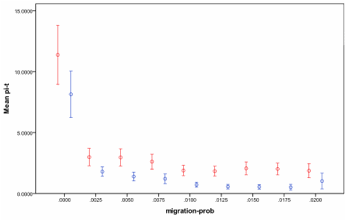
Fig. 3. Mean diversity for different migration rates with (blue) and without (red) human agents (error bars indicate a 95% confidence interval).
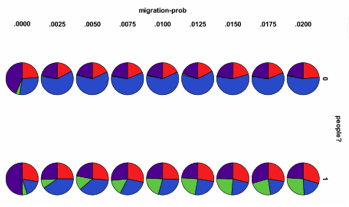
Fig. 4. Proportions of
final ecological states at final tick over independent runs for different
migration rates, with and without agents, red=plants
only, blue=with higher trophic levels, purple=monoculture, green=non
viable.
As we see from Fig. 3 and Fig. 4 above, the agents have a consistent and negative impact upon the ecologies they invade, but, generally, a higher negative impact at higher levels of migration, which tends to make the ecologies more uniform. However, as Fig. 4 indicates, this is far from a uniform effect, reducing the diversity a bit in each run. Rather it indicates an increasing proportion of ecological catastrophes (the green proportion in Fig. 4) that occur in many cases.
4. CONCLUSION
Multi-agent simulation could apply its expertise in terms of specifying
and exploring the cognitive/social abilities of agents with respect to such a
test bed, and start to tease out the complex and often counter-intuitive
effects of such abilities.
Knowledge about this could play a real part in helping us understand our
own, fragile and complex, relationship with the ecologies we inhabit and
exploit. It is time to show that
agent-technology can deliver tangible benefits to our environment and our
chances of survival.
5. ACKNOWLEDGMENTS
This research was partially supported by the
Engineering and Physical Sciences Research Council, grant number EP/H02171X/1. Many thanks to those at the Manchester
Complexity Seminar, especially Alan McKane, and those at iEMSs conference in
2012 for discussions on this subject.
6. REFERENCES
[7] Diamond, J., 2004. Collapse: How Societies
Choose to Fail or Succeed, Viking.