The Emergence of Symbiotic Groups Resulting From
Skill-Differentiation and Tags
Bruce Edmonds
Centre for
Policy Modelling,
Manchester Metropolitan University
bruce@edmonds.name
Abstract
The paper presents a evolutionary simulation where the presence of ‘tags’ and an inbuilt specialisation in terms of skills result in the development of ‘symbiotic’ sharing within groups of individuals with similar tags. It is shown that the greater the number of possible sharing occasions there are the higher the population that is able to be sustained using the same level of resources. The ‘life-cycle’ of a particular cluster of tag-groups is illustrated showing: the establishment of sharing; a focusing-in of the cluster; the exploitation of the group by a particular skill-group and the waning of the group. This simulation differs from other tag-based models in that is does not rely on either the forced donation of resources to individuals with the same tag and where the tolerance mechanism plays a significant part. These ‘symbiotic’ groups could provide the structure necessary for the true emergence of artificial societies, supporting the division of labour found in human societies.
1. Introduction
Sometimes when one is good at a certain activity one is necessarily not so good at others. That is to say that there can exist trade-offs between different abilities. In biological terms this might be the result of complex physical limitations – for example, if a species has a physique suitable for running very fast over small distances, this might limit the amount of fat its members can store to allow survival in lean times. In sociological cases this sort of trade-off might result from the amount of time that is necessary to acquire a certain skill – for example, one may not have time to learn to become a skilled musician and a skilled painter. Thus in an ecology one might have a variety of species, each of which is well adapted to exploit a different aspect of a particular environment. Similarly in our society one observes that people do not develop the same profession/skills but that there seems to be a spontaneous differentiation, so that in any locality many different skills possessed by different people are available.
When this sort of complementary differentiation is present a special kind of cooperation is possible – that where individuals with complementary skills contribute to the others any excess in what they produce/gather. In biology, when this relation has evolved into a stable relationship, this is called “symbiosis”. This sort of complementarity is very advanced in human societies; people are encouraged to specialise in terms of the skills in which they most excel, resulting in a huge range of careers and skills whose products are shared and traded in elaborate ways.
However there is a problem as to how such complementary sharing could arise in an evolutionary setting. The problem is this: from the point of view of an individual it is always better (at least in the short term) not to share the results of one’s labours but to accept those shared by others (this corresponds to the so-called “prisoner’s dilemma” (Axelrod 1984)). Thus, at any moment, those that are not sharing should do better than those who share, and hence produce more offspring. Thus it is difficult to see how groups of cooperatively sharing individuals could arise or be maintained – the so-called “tragedy of the commons” (Hardin 1968).
“Tags” are observable cues that can be used to recognise types of individuals (Holland 1993). They do not have any significant bearing on the abilities or behaviour of the individual. One can imagine a room full of people who do not know each other but are randomly given to wear different coloured badges, but who are able to exchange the badge for another if they wish. Although these badges are initially arbitrary it may (in the absence of other significant socially observable cues) allow the people to self-organise. Thus the colours may come to acquire a signifance – the significance would emerge from the social processes in the room.
There has now been a sequence of models which show that the presence of such tags can enable the evolution of a population of temporary cooperating ‘groups’ of individuals with similar tags, even when there is a possibility of being invaded by selfish indivuals who do not share or cooperate (Hales 2000, Riolo et al. 2001, Hales 2001). Basically what happens is this: a small “seed” collection of cooperative individuals with similar tags arises somehow; these out-perform the others due to their efficient cooperation and hence are grow in numbers by evolution; eventually defectors arise in the group (or invades from outside); now these defectors do even better than the others in that group and hence is preferentially reproduce until they comes to dominate that group; now the group does not do so well compared to other cooperative groups because there is little or no sharing and so the group dies. Thus what one observes is a continual rising and falling of different groups, so that in the population as a whole a relatively high level of cooperation/sharing is maintained. Clearly this depends on the facility with which new cooperative seed groups can arise compared to the speed with which established cooperative groups are infected and destroyed. This is closely linked to the rates of tag mutation compared to the rate of infection (Hales 2004).
This paper seeks to establish how tags can facilitate the development of (temporary) groups of complementary individuals in an evolutionary setting where individuals are not equipped with great cognitive abilities (to support contracts or elaborate foresight for example) and where individuals are not in any way forced to cooperate. This is important because this sort of process may allow the emergence some of the basic group infrastructure that, in turn, may facilitate the development of more sophisticated societies within an evolutionary setting. Thus the techniques and results in this paper can be seen as another step towards the full emergence of an artificial society.

Figure 1. Tag Groups in a Single Run of the Simulation: the vertical line is the tag space from 0 to 1, the horizontal axis is time and the colour indicates the number of individuals with a particular tag at a particular time.
2. Model Setup
The main assumptions that drive this model is that there are a number of different kinds of ‘nutrition’ (or ‘product’) which different individuals are specialised in gathering (or producing). However, although each individual only gathers one kind of resource they all require some of all the kinds of resource in order to survive or reproduce. Thus in order to survive and reproduce indivuals have to be given resources by other individuals that have them, otherwise they ‘starve’ and die.
Each individual has the following attributes: its special skill; a tag value; a tolerance value; and the amount of resources it has of the various kinds. The skill determines which kind of nutrition it can harvest from the environment. The tag value is an arbitrary real value in [0, 1], as is the tolerance value. The resources are a record of the amounts of each kind of nutrition they have. The tag value is the only thing that is observable by other individuals.
There is no physical space in the model, only a (one-dimensional) social ‘space’ determined by the similarity (or otherwise) of the individual’s tags. Thus one can imagine that the model represents one location or niche which they all inhabit. Each time period each individual: gathers its share of the resource it is specialised in and adds this to its store; is randomly ‘paired’ with a number of other individuals – if the difference in tag values is strictly less than its tolerance value and it has an excess in any of its resource stores it gives then a share of its resource; all individuals are ‘taxed’ a certain amount from all stores to represent consumption; finally individuals survive, die or reproduce depending upon the state of their stores. Resources degrade on transfer – thus the value of resources received in a donation event is only 0.95 of what is given.
Each time period there is a probability that the tag and/or tolerance values are mutated by the addition of Gaussian random noise. Also a small number of new individuals are continually added to the population (arriving from elsewhere). At creation, individuals are given some (low level of) initial resources. If an individual reaches a certain age or one of its resources falls to zero that individual dies. If an individual has a certain minimum in all of its resource stores it will reproduce (once in that cycle) – the initial stores in the offspring are taken from those of the parent. Individual’s only donate if their resource level reaches a minimum, which is higher that the minimum level necessary for reproduction. Thus individuals continually appear (arrive or are born), donate, consume resources, (possibly) reproduce, and die (of starvation or old age). The population level is thus variable – determined by the available resources and the efficiency of the sharing.
3. General Results
The rising and falling
of tag groups is illustrated in Figure 1.
At the start a number of tag groups form but one gains initial dominance. This dominant group then loses out to another
group between generation 200 and 300.
This is, in turn, supplanted by the group at the bottom between generation
300 and 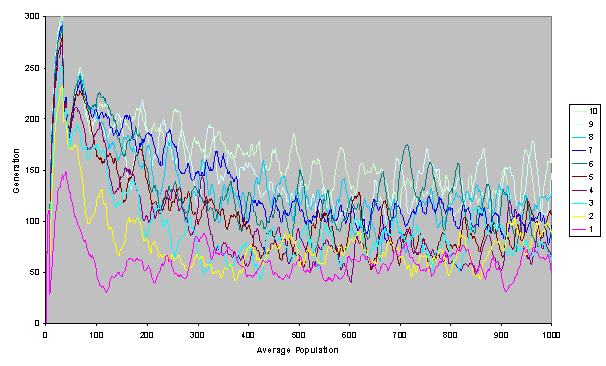
Figure 2. (average) Population with different numbers of pairing
400. Interestingly this group at the bottom seems to have seeded other groups near to it, but the whole cluster fails around generation 550, allowing a new clutch of groups to arise (towards the top).
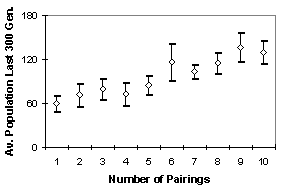
Figure 3. Average Population over last 300 generations against Number of Pairings (diamond is the average, bars are one standard deviation each way)
Figure 2 shows the population levels over runs with different levels of pairing (from 1 to 10). Each line is the average of 6 runs. One can see that the greater the number of pairings the greater the population that can be sustained with the same input resources. A summary of the over-time averages for the last 300 cycles (when the initial effects have worn off) are shown in Figure 3. This is because a higher rate of pairing there is a better chance of being paired with an individual which has a similar tag, allowing the evolution of more directed giving.
Since the population is directly related to the birth rate (which depends on individuals being given resources in nutrition types they can not themselves collect) and early death by starvation (which occurs when they are not given all the kinds they need) this indicates that effective sharing is occurring.
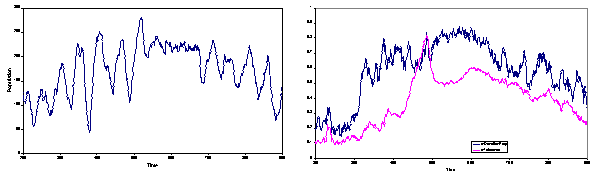
Figure 4. (left) the population against time (cycle 250 to 950); (right blue) the donation rate (from 0 to 1); (right pink) the average tolerance (from 0 to 1)
4. Case Study: The Life Cycle of a Particular Symbiotic Group
To show the resource-sharing, tag-group mechanism in detail I examine the development and demise of symbiosis in a set of groups.
The example I examine is the period between cycle 250 and 950 in a particular run (with the default parameters given in the Appendix). This period covers a cycle from low population level up to a high level and back down again (left hand panel of Figure 4). During this period the donation rate rises to a peak before falling down again (blue line, right hand panel of Figure 4) as does the tolerance level (pink line, right hand panel of Figure 4).
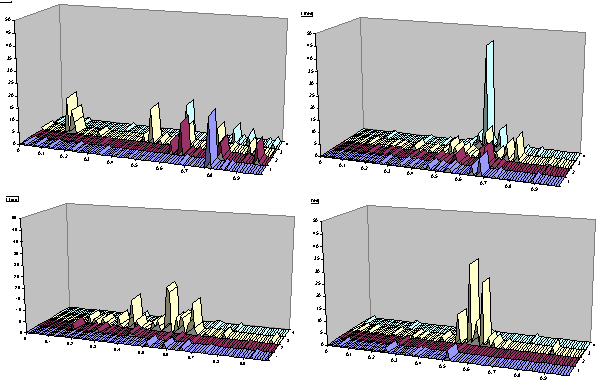
Figure 5. Life cycle of a symbiotic group: (top left, time 550) high tolerance and broad sharing; (top right, time 650) low tolerance, a tighter group, and sharing; (bottom left, time 750) medium tolerance, one skill type exploiting others; (bottom right, time 900)
Figure 5 shows a series of four ‘snapshots’ of the population distribution at cycles: 550, 650, 750 and 950. These show the distribution of tag values for each of the four skill types. By cycle 550 (top left panel of Figure 5) there has developed a scattering of tag peaks in the different skill areas, which share resources due to the high tolerances that exist at this point. By cycle 650 (top right panel of Figure 5) the group has ‘contracted’ to a tighter bunch of skill clusters with lower tolerances; by this stage one of the skills dominates the others in terms of numbers. By cycle 750 (bottom left panel of Figure 5) the sharing has become one-sided with one skill group exploiting the others, this gradually contracts to the situation at cycle 950 where this dominant group has contracted to increasingly lower tolerances. After this, these clusters dies out and a new set of related skill groups arise. This is a slightly simplified account because within this ‘life-cycle’ there are sub-cycles of groups with same skill rising and fading.
5. Related Work
There are a number of models showing how tags can facilitate the emergence of cooperation between groups of individuals with similar tags. This model is different in: (a) no individuals are forced (by the model design) to cooperate with individuals with identical tags; (b) the tolerance mechanisms whereby the range of difference which is tolerated within groups is necessary and active; and (c) there is no ‘magic’ increase in the value of donated resources from donor to recipient.
The model presented here follows that of Riolo et al. (2001), in that it uses for a tag the intensity of a single continuous variable. Tag comparisons are thus a simple matter of taking the absolute difference in tag values. This eases the display (and hence analysis) of the distributions of values that result, also in many tag models, whether one uses a continuous space of tag values, or a sufficiently large binary space seems not to make significant difference to the results. However as (Roberts and Sherratt 2002, Edmonds and Hales 2003b) showed this model relies upon the fact that individuals are forced to donate to others with an identical tag, and that the tolerance does not play any significant part[1].
Takahashi (2000) (and the tag-based variants discussed in (Edmonds and Hales 2003a)) concerned themselves with a model of generalised exchange where resource sharing resulted, but these outcomes depend on the fact that the value of a donation to a recipient is greater than the cost to a donor. That is to say that every donation has the result of increasing the resources available to the model. There is a possible (but rather forced) interpretation of this, that somehow the resource is more useful to the recipient than the donor, which could be for a variety of reasons (e.g. it was excess to the donor’s needs), but this increase in value occurs regardless of the. The model in this paper can be seen as an attempt to provide a coherent story behind the difference in value, by specifying different resource needs.
6. Towards the Emergence of Complex Artificial Societies
What has been described above shows how, in a certain setting, cooperative groups of individuals with similar tags can come into being, persist for a while and dwindle out of existence. This provides some of the ‘group infrastructure’ for more complex social structure to develop. However in order for this to occur more is needed. Essentially the groups need to be able to spawn new groups with characteristics that are similar to that of the original group, before they are infected with defectors. If, in addition to this process, the characteristics that are transmitted from group to group were potentially complex, then all the conditions necessary for the evolution of groups would be present. Presumably those groups that were more successful at resisting infection by defectors and at ‘seeding’ new groups with similar characteristics as itself would (under suitable conditions) would be more successful at seeding new groups, thus allowing for a continual process of selection and reproduction of groups that are identifiable entities in their own right (identifiable via the tags). Although evolution continues to act at the individual level, the fitness of each individual depends crucially upon the state of the group it is a member of, so if it were also the case that seeded groups had the characteristics of the groups they were seeded from (carried by the individuals who migrated out of the original group) then it would be meaningful to talk of ‘group selection’[2].
Such a process would accord with the social intelligence hypothesis (Kummer et al. 1997) and that group cultures are highly adapted to the immediate environment they inhabit (Reader 1988). The social intelligence hypothesis posits that the success of our species results more as a result of our social abilities rather than our intellectual abilities. In particular it includes a sophisticated ability to imitate others, so that skills suitable in a certain environment can spread throughout a population. This suggests that our survival may have depended upon the fact that we have socially adapted as groups to inhabit a large variety of different ecological niches, such as the Tundra and the Kalahari. The cultures developed in different groups and passed down culturally within and throughout those groups are responsible for their members ability to survive and reproduce. This model can be seen as a step forward to capturing the development of such cultural plasticity[3].
Acknowledgements
Thanks to David Hales, who has done the lion’s share of work with tags, work in which I have played but a small part. Also thanks to the many others with whom I have discussed these and similar ideas but are too numerous to mention, but including participants of MABS, M2M, ESOA and ESSA gatherings.
References
Axelrod, R. The Evolution of Cooperation, Basic Books, New York, 1984.
Edmonds, B. and Hales, D. Computational Simulation as Theoretical Experiment, CPM report 03-106, MMU, Manchester, M1 3GH, UK. (http://cfpm.org/cpmrep106.html). 2003a.
Edmonds, B. and Hales, D. Replication, Replication and Replication - Some Hard Lessons from Model Alignment. Journal of Artificial Societies and Social Simulation 6(4). http://jasss.soc.surrey.ac.uk/6/4/11, 2003b.
Hales, D. Cooperation without Space or Memory: Tags, Groups and the Prisoner's Dilemma. In Moss & Davidsson, (eds.) Multi-Agent-Based Simulation. LNAI 1979:157-166. Springer. Berlin, 2000.
Hales, D. Understanding Tag Systems by Comparing Tag Models. Second Model-to-Model Workshop (M2M2) at the Second European Social Simulation Association Conference (ESSA'04) Valladolid, Spain 16-19th Sept 2004.
Hales, D. Tag Based Co-operation in Artificial Societies. PhD Thesis, Department of Computer Science, University of Essex, UK, 2001.
Hardin, G. The Tragedy of the Commons, Science, 162:1243-1248, 1969.
Holland, J. The Effect of Labels (Tags) on Social Interactions. SFI Working Paper 93-10-064. Santa Fe Institute, Santa Fe, NM. http://www. santafe.edu/sfi/publications/wplist/1993, 1993.
Kummer, H., Daston, L., Gigerenzer, G. & Silk, J. The social intelligence hypothesis. In Weingart et. al (eds.), Human by Nature: between biology and the social sciences. Hillsdale, NJ: Lawrence Erlbaum Associates, 157-179, 1997.
Margulis, L. and Sagan, D. Microcosmos: Four Billion Years
of Evolution from Our Microbial Ancestors. Summit Books, New York, 1986.
Reader, J. Man on Earth. London: Collins, 1988.
Riolo, R. L., Cohen, M. D. and Axelrod, R. Evolution of cooperation without reciprocity. Nature, 411:441-443, 2001.
Roberts, G. and Sherratt, T. Does similarity breed cooperation? Nature 418, 499-500, 2002.
Sigmund, K. and Nowak, M. A. Evolution - Tides of tolerance. Nature 414:403, 2001.
Takahashi, N. The Emergence of Generalized Exchange”. American Journal of Sociology 105:1105-1134, 2000.
Barkow, J. H., Cosmides, L. & Tooby, J. (Eds.), The adapted mind: evolutionary psychology and the generation of culture. Oxford University Press, 1992.
Wilson, D.S. & Sober, E. Reintroducing group selection to the human behavioral sciences. Behavioral and Brain Sciences 17 (4): 585-654, 1994.
Appendix – More about the model
Sources
The model structure of tags and tolerances in a [0,1] range comes from (Riolo et al. 2000). The motivation for improving on this model came from (Edmonds and Hales 2003a, 2003b). The nutrition structure that was added on was suggested by reading texts on evolution and symbiosis, including (Margulis and Sagan 1986).
Structure
There are fixed number of nutrition types and corresponding skills for gathering that type.
There is a variable population of individuals, each of which is characterised by the following characteristics: a tag value (a real from [0,1]); a tolerance value (a real from [0,1]); a skill type (an integer); for each nutrition type: a reservoir holding an amount of that resource (a real).
Resource Flow
![]()

Resources are broadly conserved within each nutrition type. It enters via distribution and leaves via
dissipation, waste and with the death of individuals.
They principly enter the model via the direct distribution of units in the form of the different nutrition types. These are randomly distributed to these four kinds, then all those individuals who posses the appropriate skill to gather that resource kind, equally share that resource.
Also new individuals (the initial population, the 2 new individuals that enter the population each time, and the progeny of individuals that reproduce) are given a fixed amount in each reservoir (intialFood). In the case of reproduction these amounts are subtracted from the corresponding reservoirs of the parent.
Each individual is now randomly paired with a fixed number (numPairings) of other individuals. In each pairing event an amount of the resource may be transferred from giver to recipient, if some conditions are satisfied. These conditions are: (1) The recipient must be one of those randomly chosen that time; (2) the difference in tag values must be strictly less than the tolerance of the giver; and (3) the giver must have more than a set amount (foodOfTypeAboveWhichIsExtra) in the corresponding reservoir. Each donation donationCost is subtracted from the giver but only donationBenefit given to the recipient. The excess in the reservoir is shared equally among all recipients who qualify.
The individuals’ reservoirs can only store up to a fixed maximum (maxResevoir). Above that resources are simply lost.
Each unit of time, a ‘life tax’ is subtracted from each reservoir of each individual.
If an individual has accumulated more than a fixed amount (foodOfTypeNecessaryForReproduction) in all of their reservoirs then the reproduce. The resources in the offspring are subtracted from the parent.
If an individual has less than a fixed amount (foodOfTypeBelowWhichTagDies) in any reservoir then it dies, also if it has reached its maximum age (maxTagAge). Resources of those that die are lost.
Default parameter values
initialPopSize = 100
maxTime = 1000
maxRun = 1
numPairings = 6
probMutVal = 0.1
sdMut = 0.1
maxNumNew = 2
donationCost = 1
donationBenefit = 0.95
numFood = 350
numSkillBits = 2
numNutritionBits = 2
maxTagAge = 30
maxStartAge = 0
intialFood = 1
foodOfTypeNecessaryForReproduction = 4
foodOfTypeBelowWhichTagDies = 0
foodOfTypeAboveWhichIsExtra = 5
foodUsageRate = 0.25
maxResevoir = 20
maxTol = 1